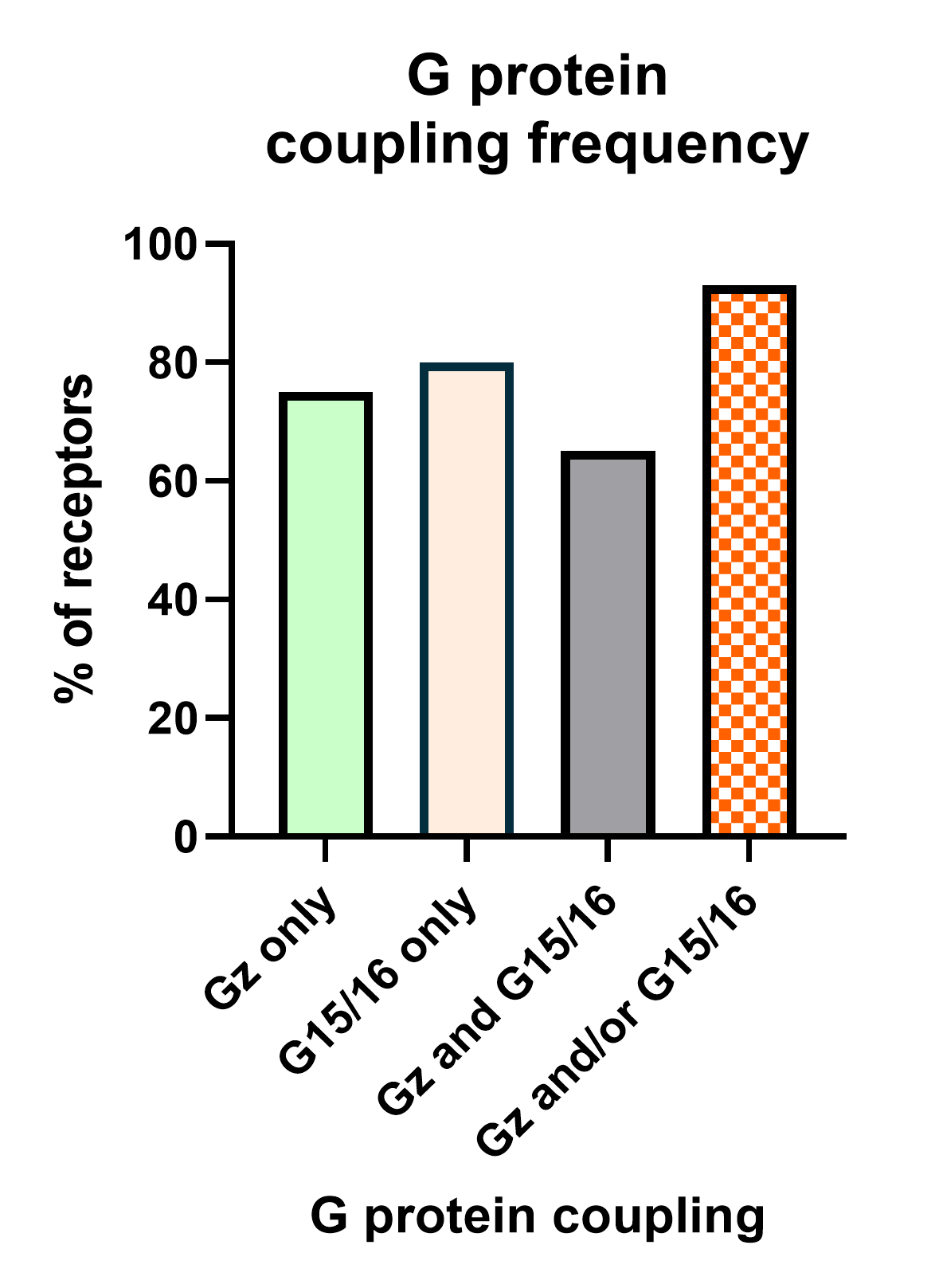Comment les biosenseurs de RCPG par BRET peuvent-ils favoriser la découverte de nouveaux médicaments?
La technologie biosens-All® de Domain Thérapeutiques repose sur des essais par biosenseurs fondés sur l’ebBRET (transfert d’énergie par résonance de bioluminescence à proximité améliorée) et codés génétiquement pour caractériser les voies de signalisation de récepteurs non-modifiés.
Nos technologies ont le même objectif sous-jacent : favoriser la découverte de nouveaux médicaments en jetant un nouvel éclairage sur les voies de transduction des signaux proximaux des récepteurs transmembranaires, sans agir sur la structure de ces récepteurs et effecteurs.
Ces deux aspects ont une importance cruciale pour qui veut concevoir la prochaine génération d’agents thérapeutiques à la lumière de connaissances approfondies sur les phénomènes intracellulaires propres aux récepteurs.
La plateforme bioSens-All® de capteurs par ebBRET, qui sert à l’étude des Récepteurs Couplés aux Protéines G (RCPG), est au cœur de la pépinière technologique de Domain Thérapeutiques.
De l’importance des RCPG
Les RCPG, dont le nombre dépasse les 800, forment la plus grande famille de récepteurs à la surface des cellules du génome humain. Comme ils interviennent dans presque tous les processus physiologiques et dans de nombreux processus pathologiques, les RCPG sont des cibles thérapeutiques majeures.
Selon les estimations, 34 % de tous les médicaments homologués par la FDA agissent sur cette famille de récepteurs. Or seulement 16 % de tous les RCPG ont été ciblés avec succès, ce qui laisse une mine de possibilités inexploitées au sein de cette famille de récepteurs.
Chez Domain Therapeutiques North America, nous nous sommes donné la mission de favoriser la découverte de nouveaux médicaments ciblant les RCPG en étudiant la pluridimensionnalité de la transduction des signaux de ces récepteurs au moyen de notre plateforme technologique bioSens-All®.
À propos de la technologie bioSens-All® appliquée aux RCPG
La plateforme bioSens-All® permet aux équipes de recherche pharmacologique d’étudier la signalisation proximale directe touchant les principaux effecteurs (protéines G et β-arrestines) mobilisés après l’activation des RCPG. La plateforme bioSens All® permet de mieux cerner les profils de signalisation proximale en fournissant une lecture fonctionnelle visant à évaluer l’activité de petites molécules ou d’anticorps à partir de données sur la signalisation qui sont pertinents sur le plan physiologique.
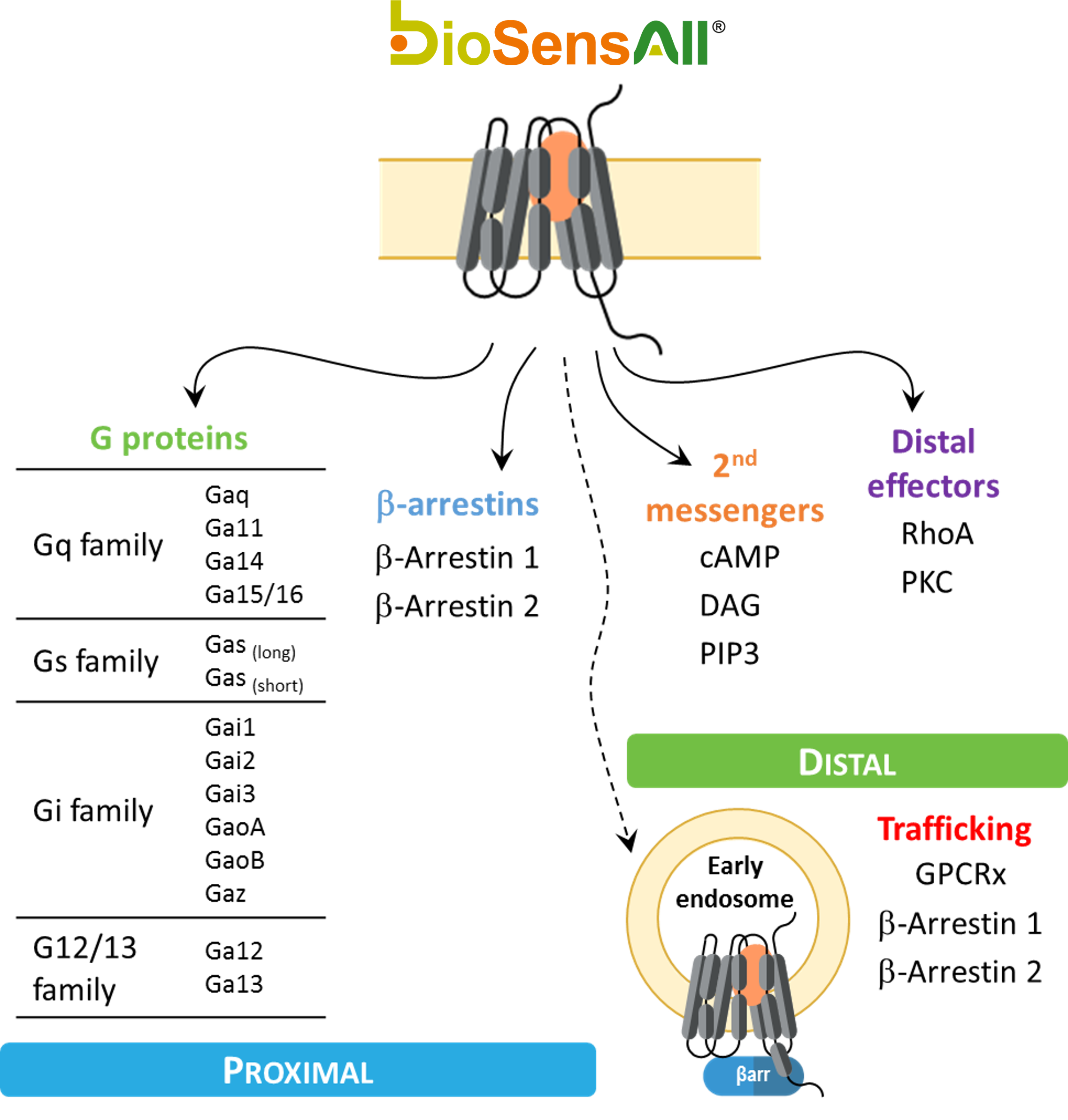
Notre objectif : exploiter tout le potentiel des RCPG dans la recherche de nouveaux médicaments grâce à un ensemble complet de biosenseurs fondés sur l’ebBRET (figure 1).
Figure 1: Panel de biosenseurs RCPG
L’importance de connaître la signature signalétique d’un ligand
Une fois activé par un ligand, un RCPG déclenche diverses voies de signalisation selon un mode propre au ligand en question, révélant ainsi une « signature signalétique».
Contrairement aux mesures distales classiques (AMPc ou Ca2+) utilisées pour déterminer la réaction d’un récepteur à un ligand donné, notre panel de biosenseurs par ebBRET jette un nouvel éclairage sur la transduction des signaux proximaux des récepteurs après l’activation des RCPG.
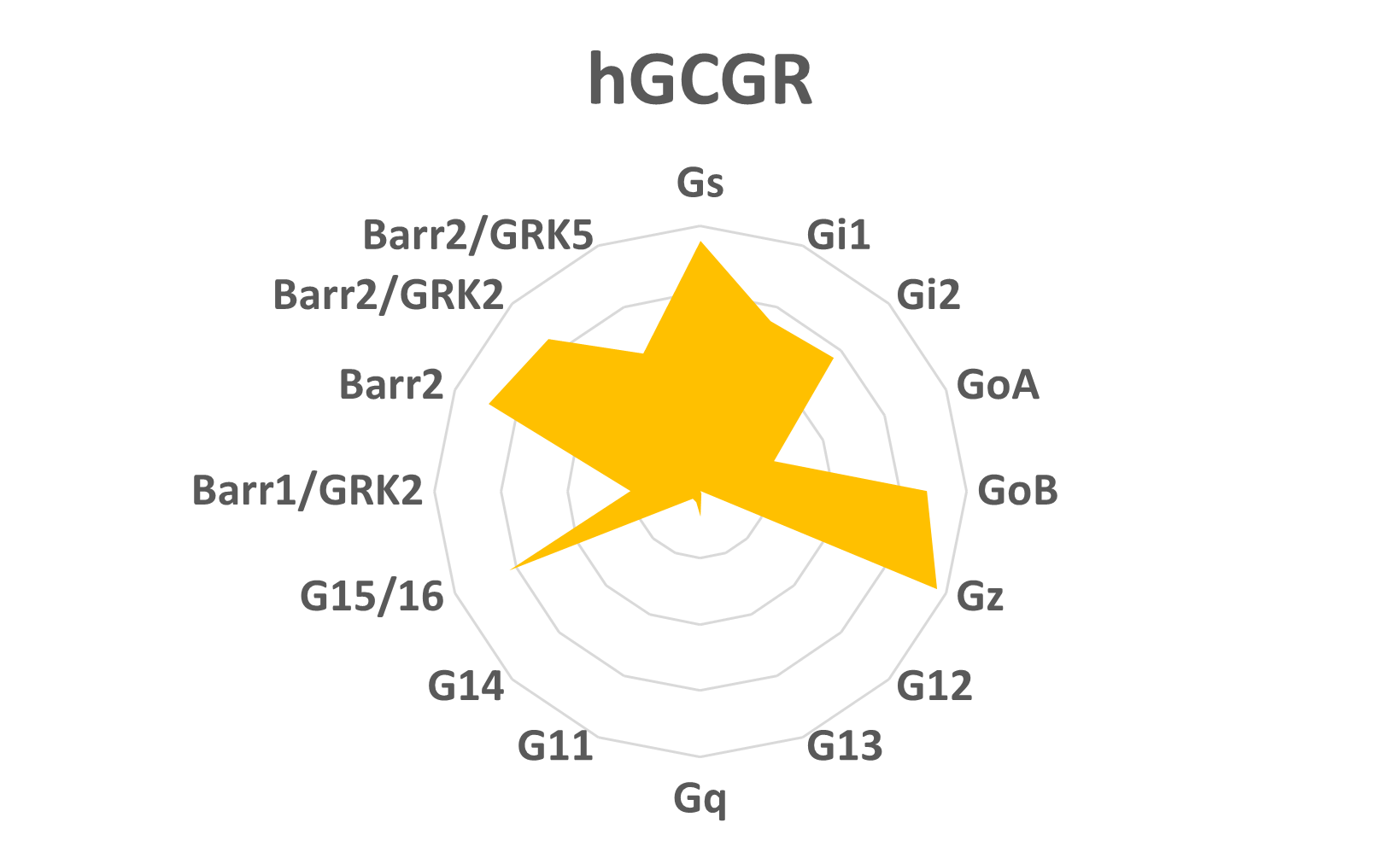
Cette information peut être utilisée pour donner le coup d’envoi à une opération de criblage visant à repérer un ligand ou groupe de ligands dotés d’une signature de signalétique particulière (figure 2) produisant les effets in vivo les plus sûrs et les plus efficaces à la lumière que ce que nous savons des effets biologiques déclenchés par l’activation de certaines voies cellulaires. La technologie bioSens-All® vise à guider les scientifiques dans l’établissement de profils complets de leurs RCPG d’intérêt et à soutenir leur programme de recherche de nouveaux médicaments.
Figure 2: Signature signalétique du ligand hGCGR
Des profils signalétiques en lien avec les effets cliniques
La technologie bioSens-All® a été utilisée pour établir des corrélations entre les signaux proximaux des récepteurs au niveau de la membrane cellulaire d’une part et les effets biologiques observés in vivo et les effets cliniques indésirables d’autre part. Plus important encore, des ligands qui n’activent pas des voies de signalisation associées à une issue clinique négative ont été caractérisés avec succès, par exemple avec le récepteur à la Niacine (HCAR2, GRP109a).
Figure 3: Les signature signalétiques corrélés aux effets in vivo
L’importance d’un système natif qui préserve la structure des RCPG
C’est un fait établi que le changement d’un seul acide aminé suffit pour changer la biologie d’un récepteur, en créant parfois une perte de fonction partielle ou totale de plusieurs voies de signalisation. Un des traits majeurs et fortement distinctifs de la technologie bioSens-All® réside dans le fait que ni les RCPG ni leurs effecteurs directs ne sont modifiés ou marqués (sauf pour les biosenseurs de Gα et de β-arrestines), ce qui fait en sorte que les récepteurs sont exprimés conformément à leur conformation physiologique et que les voies de signalisation activées ne sont altérées par aucun changement structurel, quel qu’il soit.
De plus, la possibilité de travailler avec des protéines G et des récepteurs intacts, à laquelle s’ajoute la sensibilité de notre technologie par ebBRET, comporte de nombreux avantages. Premièrement, cela permet de détecter des signaux émis par des RCPG de façon endogène dans des lignées cellulaires pertinentes sur le plan physiologique. Cela permet également de déceler l’activation de protéines G avec des composés natifs dans les cellules exprimant les récepteurs et les protéines G endogènes d’intérêt en proportion suffisante. Il est donc possible de s’approcher encore plus de conditions physiologiques inaltérées en ne surexprimant pas le récepteur, c’est-à-dire en travaillant avec des cellules exprimant le RCPG d’intérêt de façon naturelle, pour étudier son comportement biologique dans un contexte correspondant le plus possible à sa physiologie.
Enfin, la grande sensibilité de notre système peut être mise à profit pour déceler des agonistes inverses en mesurant le niveau d’activité constitutive du récepteur et sa réversibilité potentielle par un ligand donné.
Dans l’ensemble, le fait que cette technologie ne comporte pas de marquage du RCPG et des effecteurs est un avantage crucial pour tirer des conclusions fiables et étoffées à partir de ces données sur la signalisation.
Des biosenseurs fondés sur le BRET pour concevoir la prochaine génération d’agents thérapeutiques
La connaissance de la signature signalétique d’un ligand, qu’il s’agisse d’une petite molécule ou d’un agent biologique, peut réellement faire une différence pour la découverte et développement de nouveaux médicaments.
La qualité de l’information fournie par la technologie bioSens-All® peut être un atout majeur à différents niveaux :
Analyse spatiotemporelle et de localisation en temps réel
Tous nos biosenseurs par ebBRET peuvent être suivis en temps réel et dans divers compartiments intracellulaires. Cette caractéristique procure un autre niveau d’analyse exploitable. Comme deux ligands différents peuvent avoir une signature signalétique semblable du point de vue des mesures en point final, mais une cinétique d’activation totalement différente se traduisant par des effets physiologiques différents, l’analyse en temps réel peut se révéler particulièrement utile pour discriminer des ligands et mieux cerner les effets fonctionnels de chacun.
Pour obtenir plus de renseignements sur l’analyse spatiotemporelle
Reconnaissance d’une signalisation biaisée et de la signature signalétique souhaitée
Grâce à une meilleure connaissance de la signature signalétiqued’un ligand, il est possible de concevoir précisément le médicament qui permettra d’activer certaines voies de signalisation, tout en en inhibant certaines autres. Ce concept fondamental, la « signalisation biaisée », peut multiplier les chances de découvrir un nouveau médicament et amplifier le potentiel thérapeutique d’un médicament et d’un récepteur, tout en inhibant leurs effets secondaires.
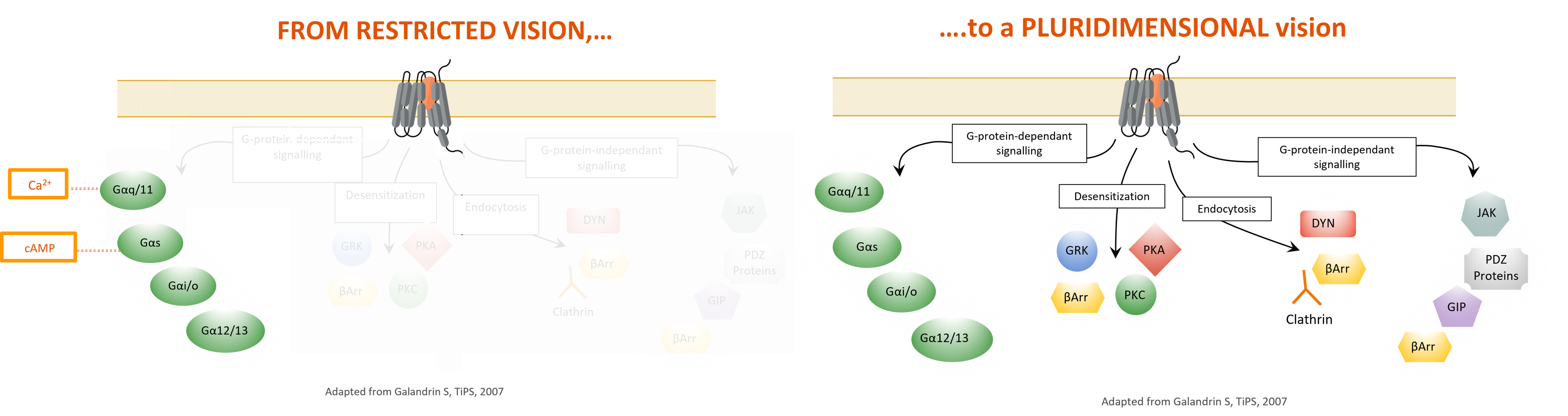
Les RCPG ont historiquement été vus comme des interrupteurs de signalisation binaires en équilibre entre le mode « allumé » ou « éteint ». Selon ce concept, les ligands sont classés en trois groupes ꟷ les agonistes, les agonistes inverses ou les antagonistes ꟷ en fonction de leur effet global sur cet équilibre (les agonistes faisant passer l’équilibre vers l’état « allumé » et les agonistes inverses le faisant passer vers l’état « éteint »). Deux grandes découvertes complémentaires survenues au cours de la dernière décennie, soit la pluridimensionnalité de l’efficacité des signaux et l’agonisme biaisé2, ont révolutionné notre compréhension des propriétés biologiques des RCPG et du comportement pharmacologique des ligands ciblant ces récepteurs. Contrairement à la vision classiquesur la signalisation des RCPG, il est aujourd’hui évident que ces récepteurs peuvent avoir plusieurs états actifs (c.-à-d. « allumé »3) et intervenir dans plusieurs cascades de signalisation (potentiellement croisées) en aval en se couplant directement à des protéines G hétérotrimériques et à des effecteurs indépendants des voies des protéines G (β-arrestines). De plus, la notion de biais s’applique aussi à un médicament donné agissant sur un récepteur et pouvant exercer une activité différente (parfois opposée) sur les diverses voies couplées au récepteur en question.
L’efficacité d’un médicament est donc considérée comme un paramètre pluridimensionnel. Dans le prolongement de cette observation se trouve la notion d’agonisme biaisé (ou sélectivité fonctionnelle) qui pose comme hypothèse que des ligands ayant une structure dissemblable peuvent stabiliser des conformations de récepteurs uniques, chacune de ces conformations se liant différemment aux effecteurs en aval. Par conséquent, les différents ligands d’un récepteur donné peuvent conférer à ce dernier des signatures signalétiqueset des propriétés biologiques distinctes. Le potentiel de l’agonisme biaisé réside dans son applicabilité au développement de ligands destinés aux RCPG qui mobilisent sélectivement certaines voies thérapeutiques tout en inhibant celles provoquant des effets nocifs ou en y restant indifférents. À terme, les ligands biaisés pourraient afficher des profils thérapeutiques supérieurs dans de nombreux domaines pathologiques et indications.
Nous démontrons ci-après comment les biosenseurs bioSens-All® ont été utilisés pour caractériser et différencier les signatures signalétiquesdes ligands du récepteur 1 de la parathormone (PTH1R) humaine endogène et de synthèse (soit la parathormone [PTH] et la Tyr1-PTH, respectivement).
Pour obtenir plus de renseignements sur la signalisation biaisée avec le PTH1R comme exemple
Différentes méthodes pour aborder les RCPG orphelins
Même si les RCPG constituent des cibles thérapeutiques privilégiées – avec34 % de tous les médicaments homologués par la FDA exercant leur action sur cette famille de récepteurs –, il n’en demeure pas moins que 16 % seulement de tous les RCPG sont ciblés.
Plus de 100 RCPG sont encore orphelins, c’est-à-dire qu’on ne leur connaît aucun ligand à ce jour. Les traits pharmacologiques et de signalisation de ces RCPG restent donc hors de portée, puisqu’aucun modulateur endogène ou exogène n’a encore été découvert.
La technologie bioSens-All® peut être utile pour aborder les RCPG orphelins de deux façons distinctes :
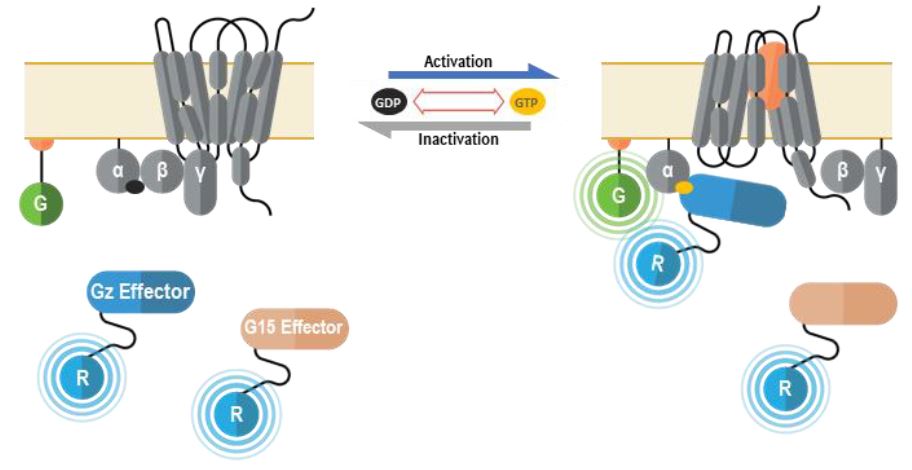
La première réside en la capacité de cette technologie de détecter des liaisons effectrices indépendantes de ligands, ou constitutives. L’activité constitutive est aujourd’hui largement reconnue comme une caractéristique intrinsèque des RCPG, qui a également été objectivée lors d’études in vivo (Damian et al., 2012; Corder et al., 2013). Une telle activité constitutive suivant la surexpression d’un RCPG peut servir à repérer la voie effectrice intervenant dans les signaux d’un récepteur orphelin donné, un composé agoniste devenant du coup moins nécessaire. Selon cette stratégie, un récepteur est surexprimé à divers degrés et des mesures sont obtenues pour de nombreuses voies, comme le montre la figure ci-dessous.
La deuxième stratégie consiste à combiner nos biosenseurs de sous-unités Gz et G15.
Nous avons récemment terminé l’analyse des liaisons aux protéines G de 100 RCPG. Or cet exercice a révélé que 95 % de ces récepteurs activent les sous-unités Gz et/ou G15 (figure 6). Par conséquent, la création de ce capteur capable de mesurer l’activation des deux voies simultanément a pour résultat un senseur puissant et quasi universel pouvant être utilisé à des fins de déorphanization de récepteurs orphelins.
De plus, ce capteur peut servir à réaliser des campagnes permettant de tester la sécurité biologique de composés en testant parallèlement un panel de récepteurs impliqués dans des effets toxiques ou non voulus.
Pour finir, il est possible de l’utiliser pour détecter des agonistes inverses en mesurant le niveau d’activité constitutive d’un récepteur et sa réversibilité potentielle au moyen d’un ligand donné.
BRET : principe et biosenseurs
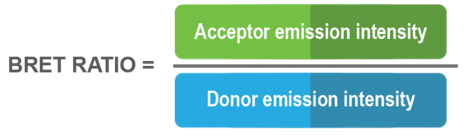
Le transfert d’énergie par résonance de bioluminescence (BRET) est un phénomène naturel reposant sur un transfert d’énergie non radiatif entre une protéine donneuse luminescente (soit une luciférase) et une protéine receveuse fluorescente (p. ex. une protéine verte fluorescente).
En présence d’oxygène, la luciférase catalyse l’oxydation de son substrat, ce qui produit de la lumière. Quand la luciférase et la protéine receveuse sont suffisamment proches dans l’espace, cette dernière absorbe l’énergie émise par la réaction entre la luciférase et son substrat, et émet à son tour un signal fluorescent. Le signal de BRET se calcule en divisant l’intensité de l’émission produite par la protéine receveuse par celle générée par la protéine donneuse :
L’importance de cette valeur radiométrique de BRET est essentiellement tributaire de l’orientation et de la proximité physique relatives entre la protéine donneuse et la protéine receveuse (plus la protéine donneuse et la protéine receveuse sont proches l’une de l’autre, plus le transfert d’énergie est efficace , plus le signal de BRET est fort).
Le BRET se produit généralement entre 1 et 10 nm (de 10 à 100 Å) de distance. Cette aire se compare aux dimensions des macromolécules biologiques et cadre bien avec la plupart des interactions biologiques, ce qui fait du BRET un système idéal pour l’étude des interactions macromoléculaires et des changements de conformation dans les cellules vivantes.
La plateforme bioSens-All® fournit des biosenseurs fondés sur le BRET pour étudier la signalisation dépendante des protéines G et celle dont la médiation est assurée par les β-arrestines en aval des récepteurs couplés aux protéines G (RCPG).
Ces biosenseurs servent à mesurer :
l’activation de 14 protéines G hétérotrimériques différentes de même que les β‑arrestines 1 et 2
la production de messagers secondaires
cAMP
diacylglycerol (DAG)
phosphatidylinositol (3,4,5)-trisphosphate (PIP3)
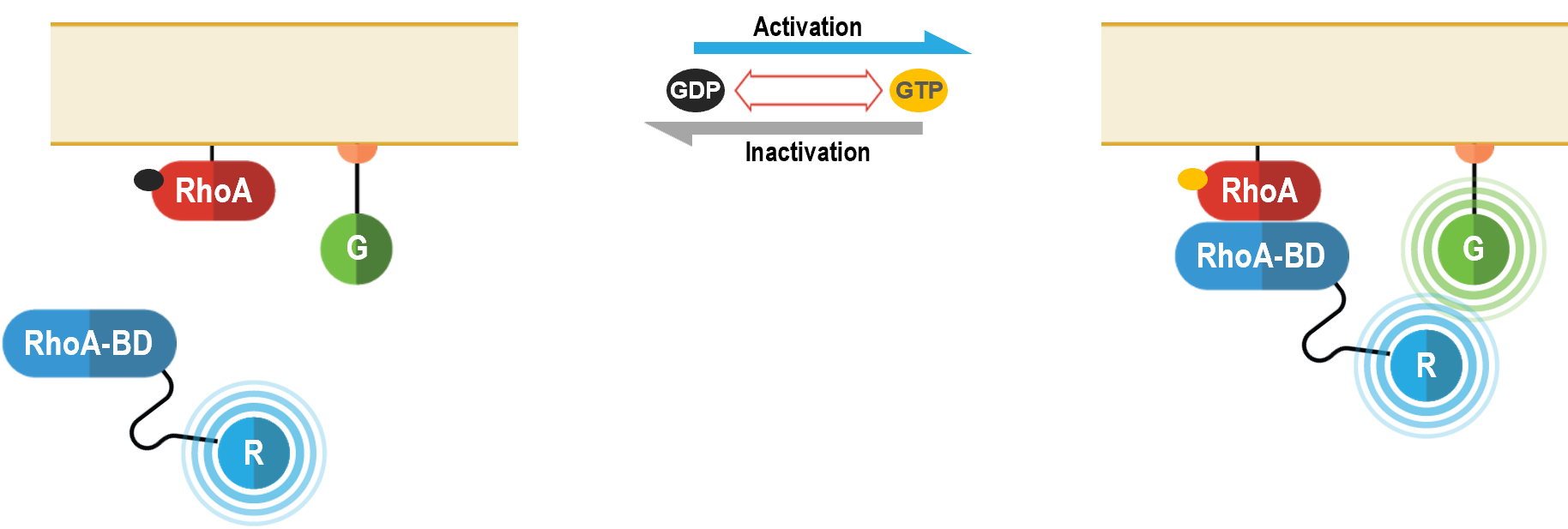
la mobilisation des protéines effectrices en aval
RhoA
Activation de PKC
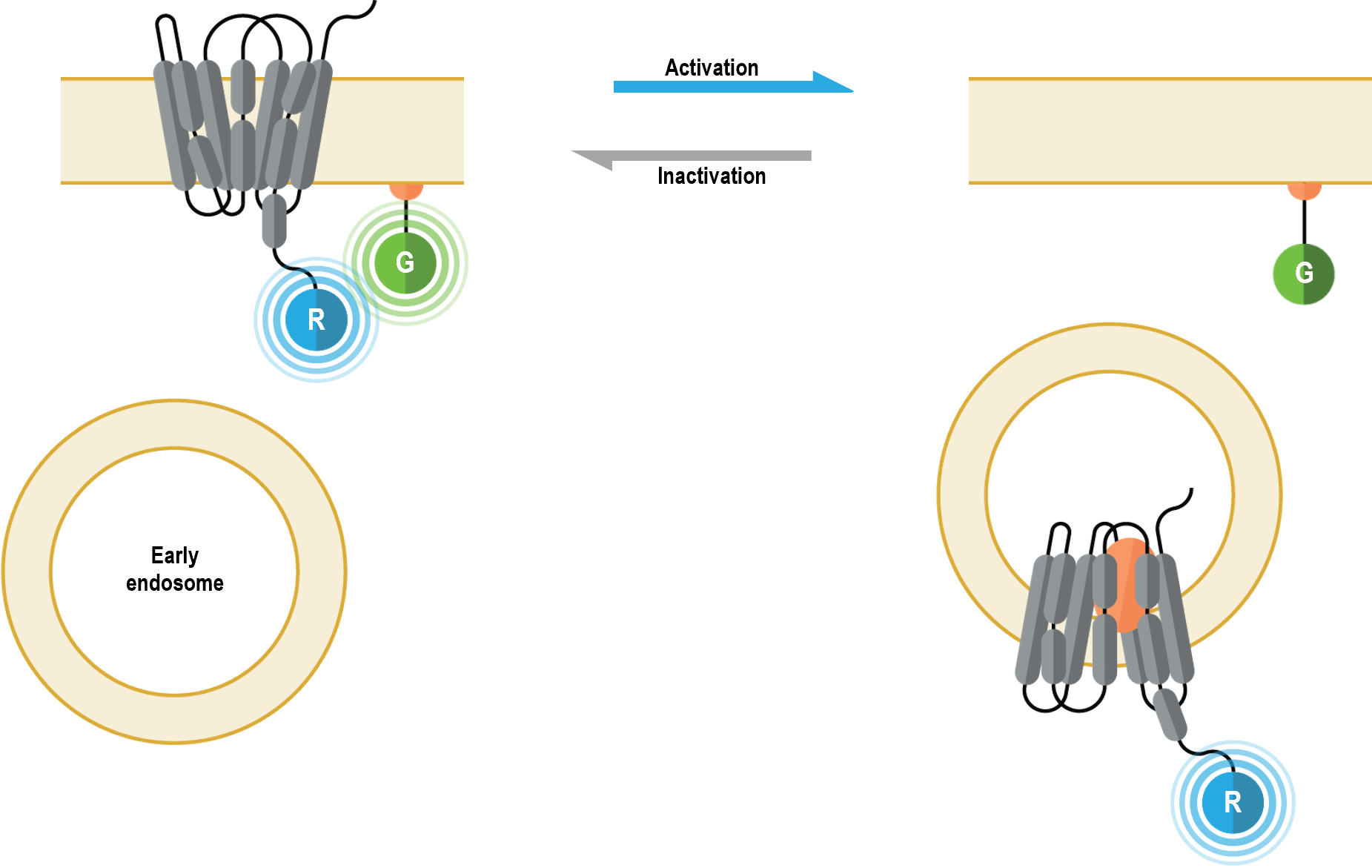
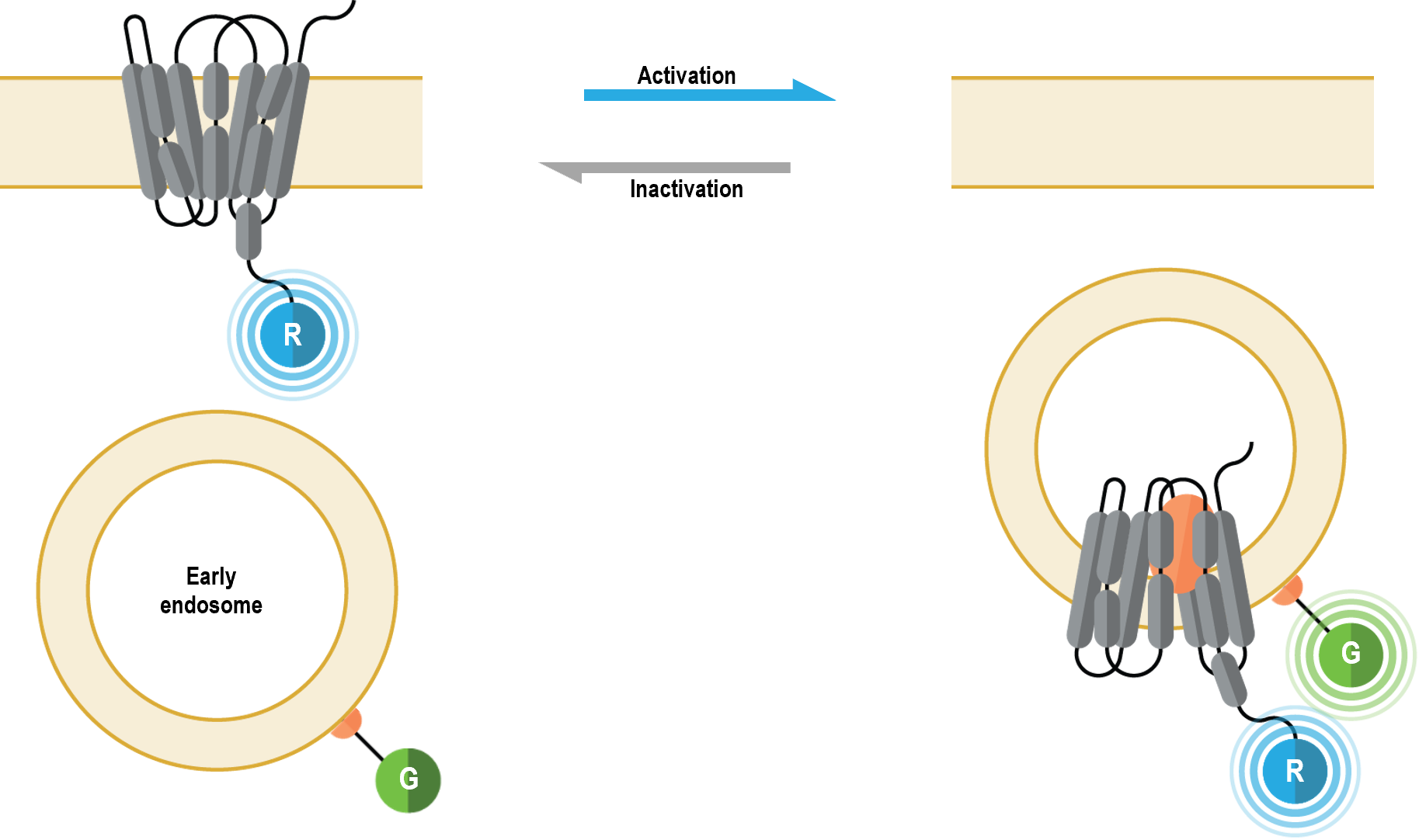
l’emplacement et le trafic des récepteurs (membrane cellulaire vs endosomes précoces)
Les biosenseurs qui sont au cœur de la plateforme bioSens-All® sont le fruit de différentes stratégies. Ils peuvent être répartis dans les catégories fonctionnelles suivantes :
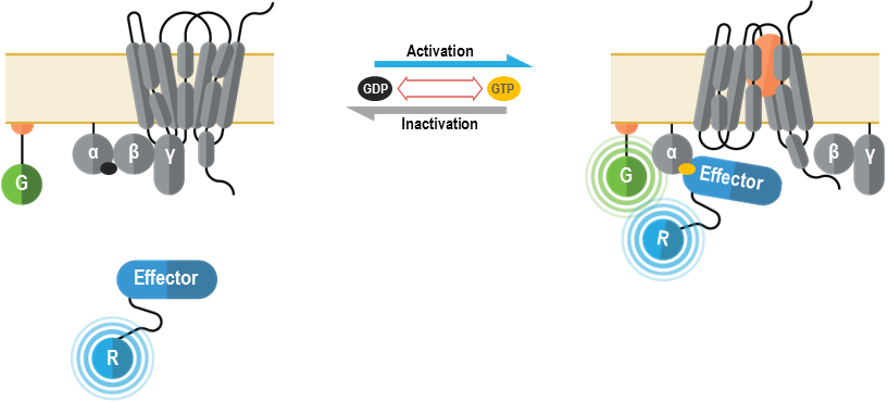
A) Biosenseurs de l’activation de protéines G hétérotrimériques
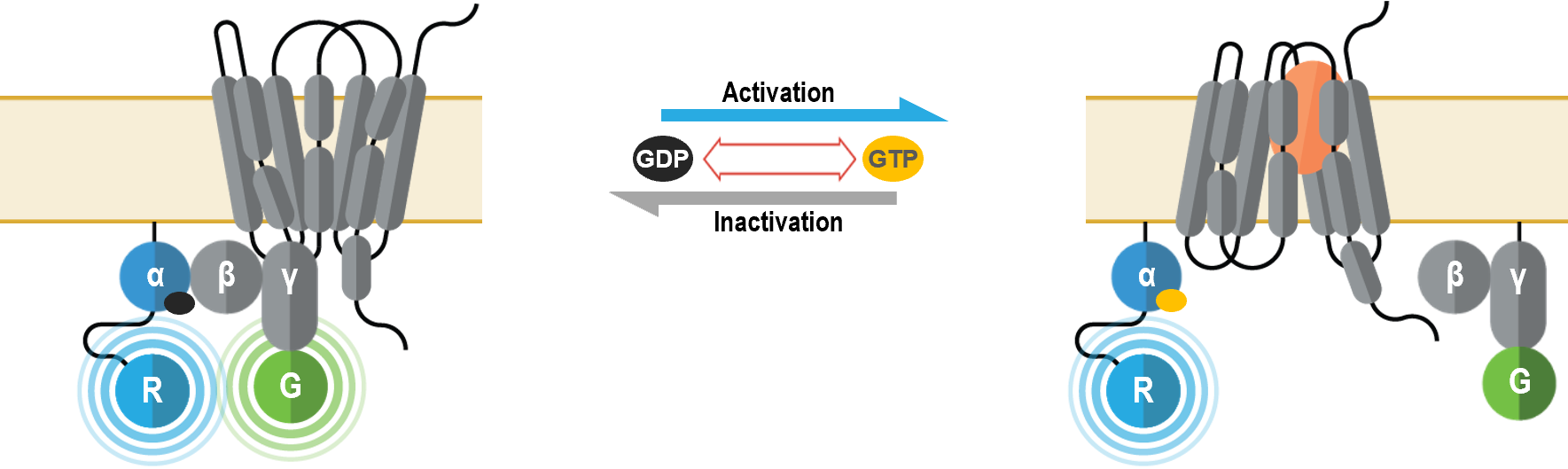
I. G Biosenseurs de l’activation de protéines G attribuable aux sous-unités Gα/Gγ (GABY)
Ces senseurs multimoléculaires ont été conçus aux fins de vérification des changements conformationnels survenant dans le complexe qu’est une protéine G hétérotrimérique après activation des sous-unités Gα et interaction avec un effecteur. Ils peuvent être employés pour mesurer l’activation spécifique de chaque sous-type de Gα.
II. Biosenseurs des sous-unités Gα membranaires (GAPL)
Ces senseurs multimoléculaires détectent le recrutement sur la membrane cellulaire d’une protéine effectrice qui interagit avec les sous-unités Gα au moment de l’activation des protéines G. Ils peuvent être utilisés pour mesurer l’activation spécifique de chaque sous-unité Gα.
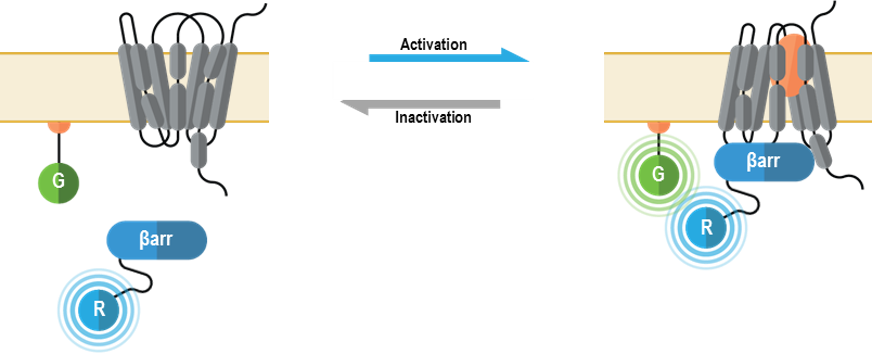
I. Biosenseurs détectant le recrutement de ß-arrestines membranaires
Ces senseurs multimoléculaires détectent le recrutement de la ß‑arrestine 1 ou de la ß‑arrestine 2 sur la membrane cellulaire après la liaison aux récepteurs.
Une stratégie semblable est offerte pour détecter le recrutement des ß-arrestines vers les endosomes précoces.
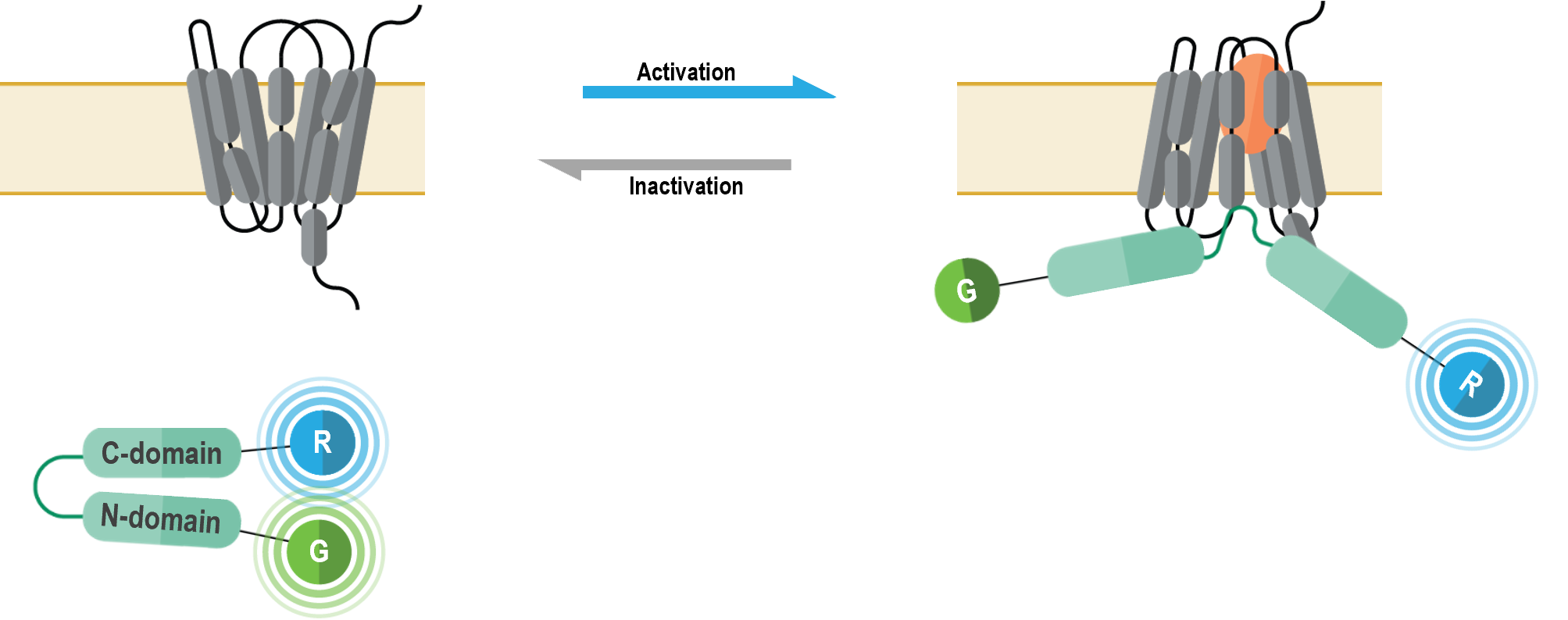
II. Beta-arrestin double brilliance conformation biosensors
Ces biosenseurs monomoléculaires sont utilisés pour observer les changements de conformation survenant dans les β-arrestines 1 et 2 au moment de l’activation des RCPG.
Les biosenseurs multimoléculaires de l’emplacement et du trafic des récepteurs permettent de suivre en temps réel l’évolution de la membrane cellulaire et l’emplacement endosomique des RCPG ou des ß-arrestines après leur activation. De plus, ces senseurs peuvent être utilisés de manière complémentaire pour étudier les caractéristiques cinétiques du trafic entre la membrane cellulaire et les endosomes.
I. Biosenseurs de localisation à la membrane plasmique
Les biosenseurs multimoléculaires de l’emplacement et du trafic des récepteurs permettent de suivre en temps réel l’évolution de la membrane plasmique et l’emplacement endosomique des RCPG après leur activation. De plus, ces capteurs peuvent être utilisés de manière complémentaire pour étudier les caractéristiques cinétiques du trafic entre la membrane plasmique et les endosomes.
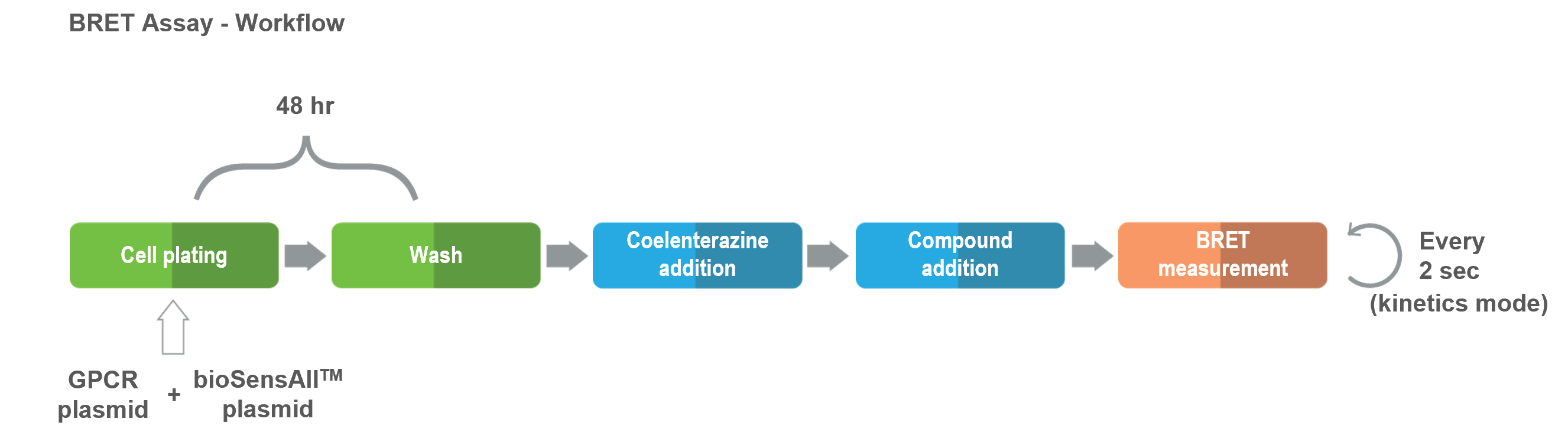
bioSens-All® est une plateforme extrêmement dynamique et polyvalente qui peut être facilement adaptée de façon à répondre à l’ensemble de vos questions et de vos besoins. Cet essaia été employé avec succès pour des ligands de nature variée (p. ex. petites molécules, peptides, agents biologiques). Toutes les applications de cette technologie sont fondées sur le protocole élémentaire suivant et sont compatibles avec des plaques de 96 et de 384 puits :
les cellules sont transfectées avec des vecteurs codant pour le RCPG d’intérêt non-marqué et le biosenseur désiré, puis sont réparties dans les microplaques de 96 ou de 384 puits;
48 heures après la transfection, les composés sont incubés avec les cellules et l’activité des biosenseurs est mesurée (par BRET).
À ce jour, plus de 100 RCPG de type naturel et plus de 14000 variants de RCPG ont été caractérisés au moyen de la technologie bioSens-All®. Les signatures signalétiques de nombreux ligands ont été définies pour de nombreux récepteurs.